Immersion in water is the closest approximation to weightlessness. Moreover, teleosts, through their invention of the gas bladder, have learned to escape the drag of gravity with the least energy expenditure. The downside of being under water is the fact that any movement has to push aside water, a particularly dense and heavy medium. The physics and physiology of fish swimming are summarized in Blake (1983) and Webb (1984) and the most recent comprehensive review in this field is that of Videler (1993).
This section describes the manner in which fish swim, something they do in a surprising number of ways; also, available records on the swimming speed of fish are presented.
The classification of the swimming modes of fish adopted here stems from Lindsey (1978), who reviewed much of the earlier literature on this topic. This classification consists of two levels, the first (Roman numerals) describing what may be called the swimming ‘type’, the second (bullets) describing the swimming ‘mode’ proper, viz.
Movements of body and/or caudal fin:
Anguilliform;
Subcarangiform;
Carangiform;
Thunniform; and
Ostraciiform.
Undulation of median or pectoral fins:
Amiiform;
Gymnotiform;
Balistiform;
Rajiform; and
Diodontiform.
Oscillations of median or pectoral fins:
Tetraodontiform;
Labriform.
The modes in (I) imply a gradual transition from undulation of the entire body (including the trunk) being used for propulsion (anguilliform mode) to propulsive forces being generated only by the oscillating caudal fin (thunniform and ostraciiform modes).
The ranking of the modes in I and II to III further implies a gradual transition from undulations to oscillations as the movements generating the major propulsive force, and thence the above scheme can also be represented as a graph, with a trunk-to-caudal-fin axis as ordinate and an undulation-to-oscillation axis as abscissa (see Fig. 1 in Lindsey 1978).
This implies that the assignment of a swimming mode to a given fish species will always contain a subjective element, even if we ignore the fact that fish may have two swimming modes (we consider here only the dominant mode, e.g., labriform in parrotfish, Family Scaridae, even if parrotfish revert to the subcarangiform mode when escaping from presumed danger (authors’ pers. field obs.)).
Thus, the entries of this table (i.e., the choices of the types and modes listed above) may be revised from time to time as a result of our familiarization with this topic and its literature. This will not, however, affect the first set of entries, based on Lindsey’s examples [Species explicitly assigned to a certain swimming mode by Lindsey (1978) in both the MainRef. and Ref. fields] and their obvious extensions (e.g., from Anguilla anguilla to all Anguillidae, and thence to all Anguilliformes).
Note also that this table presently pertains only to juvenile and adult fishes. Fish larvae¾ for obvious reasons¾ have a limited repertoire of swimming types and modes.
The aspect ratio of the caudal fin of a species closely correlates with its average level of activity (Pauly 1989). The Aspect ratio (A) of the caudal fin is calculated from:
A = h2 / s
where h is the height of the caudal fin and s its surface area (Fig. 52). The proximal border of the caudal fin surface is defined as a straight vertical line through the narrowest portion of the caudal peduncle, i.e., the portion of the peduncle surrounded by the fin is considered part of the caudal fin area (see Fig. 52).
Fig. 52. Aspect ratio (A = h2/s, h = height of the
caudal fin; s = surface area of fin) of a pelagic fish (A = 7.5) and a bottom
dweller (B = 0.6). Note the correspondence between aspect ratios and modes of
life.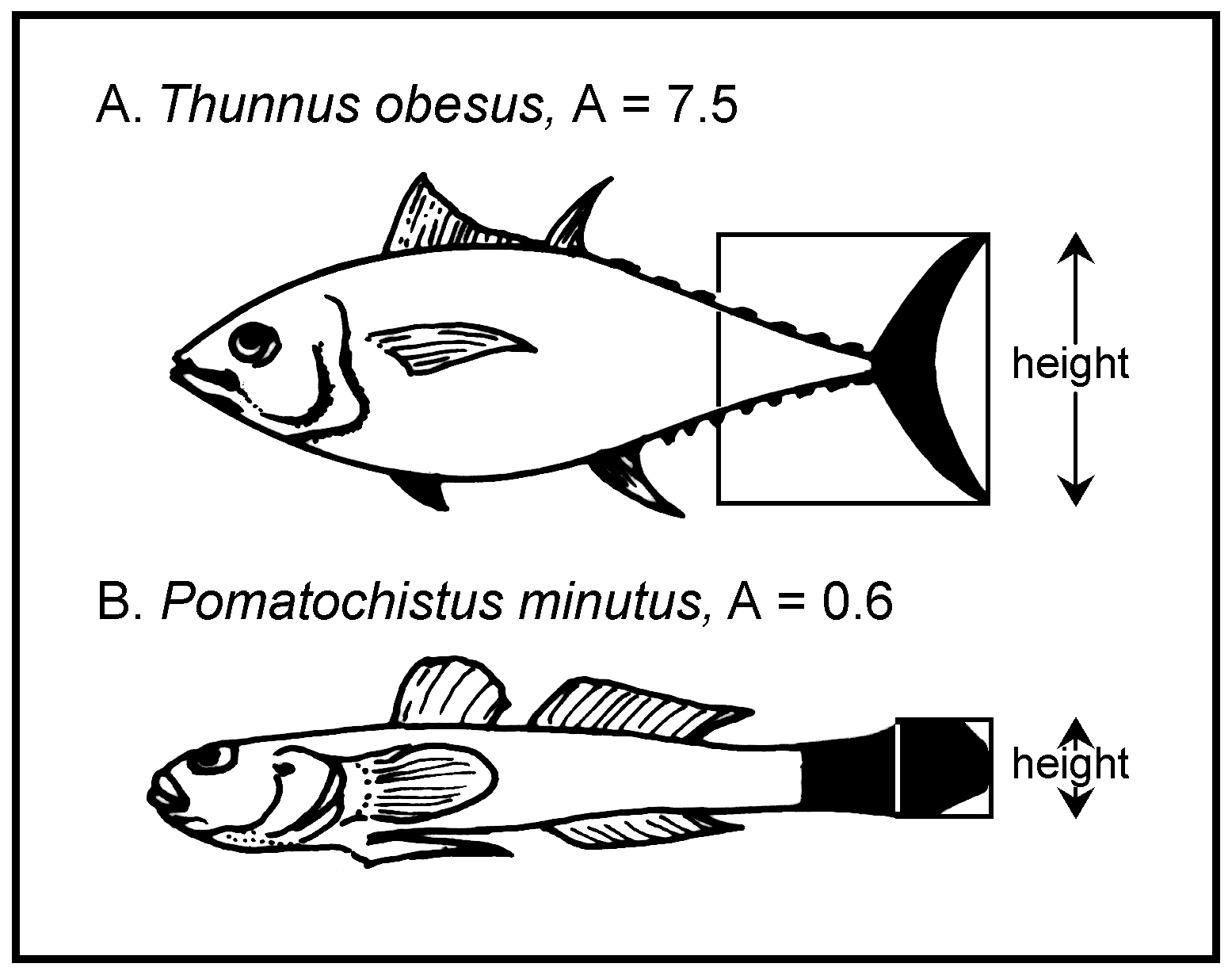
Note that this definition of A differs slightly from that presented in Pauly (1989), where only the caudal fin proper was used for calculating s.
A reference may be given to document an aspect ratio; when none is given, the aspect ratio was calculated, by planimetry, from a fish picture in FishBase or another readily available source.
The SWIMMING table contains swimming type and mode for over 2,700 species.
The SPEED table contains 255 records with maximum swimming speeds for 80 species. The information was extracted from over 50 references such as Bainbridge (1958, 1960), and Webb (1971) and compilations such as Sambilay (1990). An effort was made to distinguish between ‘sustained’ (i.e., maintained for more than 3 minutes), and ‘burst’ swimming (maintained for a few seconds only) (see Fig. 53), as well as other swimming modes (Hammer 1995).
The SPEED table consists of the following fields:
Length: This field pertains to the length of fish in cm as stated in the publication. The type of length measurement used consists of the following choices: SL (Standard Length); FL (Fork Length); TL (Total Length); BL (for the term ‘body length’, stated in the publication but without the type of length measurement being indicated).
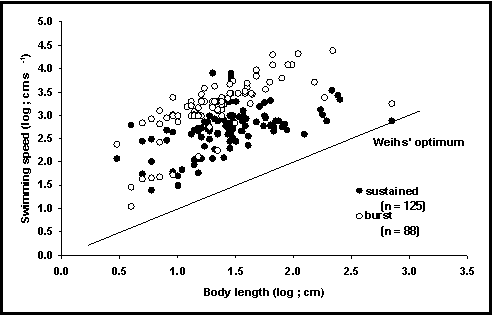
Fig. 53. Relationship between swimming speed and body length of
fishes. Note that burst speed is about 10 times higher than sustained speed.
Note also that no fish appears to swim at speeds below Weihs’ (1973) line of
minimum energy expenditure per distance covered, whose slope (1 : 1, implying
strict proportionality of speed and length), however, is the same as that of the
observations in this figure.
The Swimming speed gives the speed of the fish per second as reported in the source. A choice field is provided for the type of length measurement used. The choices are the same as above (SL, FL, TL, BL) and m/s (for meter per second). A calculated Speed field in meter per second is also provided (see Derived values).
Mode: This field describes the mode of swimming as: sustained (swimming at this speed for a prolonged time); burst (a maximum swimming speed which can be maintained for less than a minute only); other.
Comment: States the length or weight measurement, or mode of swimming if not in the choice list. Any transformation of length type should also be stated here.
Derived values: Gives the transformation values from different units of speed to standard length per second (SL/s) and to meter per second (m/s). This makes it possible to compare the swimming performance of fishes with forked, rounded or other shapes of caudal fin. Where available, the a and b values used to transform fork length or total length to standard length are given as used in:
SL = a + b × L
where L is the reported fork or total length. If the transformation is based on one measurement only, hopefully a typical adult, the intercept a is set to 0 (see also the ‘LENGTH-LENGTH table’, this vol.).
You get to the SWIMMING and SPEED tables by clicking on the Biology button in the SPECIES window, the Morphology and physiology button in the BIOLOGY window, and either the Swim. type or Swim. speed buttons in the next window.
On the Internet, you get to the SPEED table by clicking on the Speed link in the ‘More information’ section of the ‘Species Summary’ page. You can create a list of all species with available data by selecting the Swim. Speed radio button in the ‘Information by Topic’ section of the ‘Search FishBase’ page. As of December 2000, information on swimming type was not yet available on the Internet.
Bainbridge, R. 1958. The speed of swimming of fish as related to the size and to the frequency and amplitude of the tail beat. J. Exp. Biol. 35(1):109-133.
Bainbridge, R. 1960. Speed and stamina in three fish. J. Exp. Biol. 37(1):129-153.
Blake, R.W. 1983. Functional design and burst-and-coast swimming in fishes. Can. J. Zool. 61:2491-2494.
Hammer, C. 1995. Fatigue and exercise tests with fish. Comp. Biochem. Phsyiol. 112A(1):1-20.
Lindsey, C.C. 1978. Form, function, and locomotory habits in fish, p. 1-100. In W.S. Hoar and D.J. Randall (eds.) Fish physiology. Vol. II. Academic Press, New York.
Pauly, D. 1989. A simple index of metabolic level in fishes. Fishbyte 7(1):22.
Sambilay, V.C., Jr. 1990. Interrelationships between swimming speed, caudal fin aspect ratio and body length of fishes. Fishbyte 8(3):16-20.
Videler, J.J. 1993. Fish swimming. Chapman and Hall, London. 260 p.
Webb, P.W. 1971. The swimming energetics of trout. Thrust and power output at cruising speed. J. Exp. Biol. 55:489-520.
Webb, P.W. 1984. Form and function in fish swimming. Sci. Am. 251:58-68.
Weihs, D. 1973. Optimal fish cruising speed. Nature 245 (Sept. 7):48-50.
Rainer Froese, Armi Torres, Crispina Binohlan and Daniel Pauly